Animal diversity
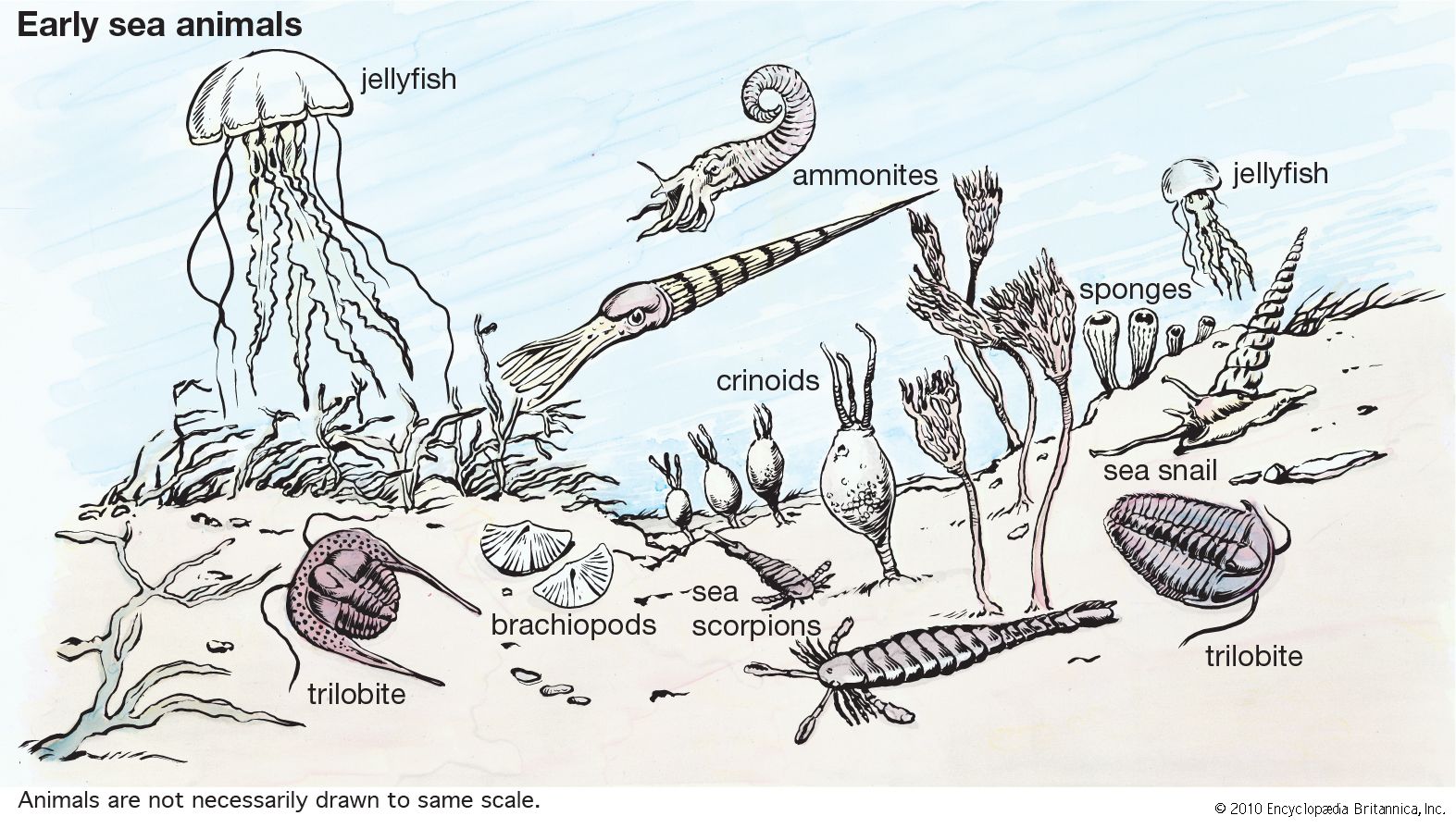
The diverse appearance of animals is mostly superficial; the bewildering variety of known forms, some truly bizarre, can be assorted among a mere half-dozen basic body plans. These plans are established during the embryonic stages of development and limit the size and complexity of the animals. Symmetry, number and relative development of tissue layers, presence and nature of body cavities, and several aspects of early development define these fundamental modes of organization.
Parazoa: a cellular level of organization

Although the two phyla in this subkingdom, Porifera (sponges) and Placozoa, lack clearly defined tissues and organs, their cells specialize and integrate their activities. Their simplicity has been adaptive, and sponges have remained important in benthic marine habitats since their origin. The sessile, filter-feeding way of life shown by sponges has favoured a body plan of radial symmetry, although some members have become asymmetrical. The shape of the creeping, flattened placozoans is irregular and changeable.
Radiata: a tissue level of organization

The two coelenterate phyla (Cnidaria and Ctenophora) advanced in complexity beyond the parazoans by developing incipient tissues—groups of cells that are integrally coordinated in the performance of a certain function. For example, coelenterates have well-defined nerve nets, and their contractile fibres, although only specialized parts of more generalized cells, are organized into discrete muscle units. Because discrete cells of different types do not carry out the internal functions of the animals, coelenterates are considered to be organized at only a tissue level.
The integration of cells into tissues, particularly those of nerve and muscle, permits a significantly larger individual body size than is possible with other modes of body movement. Flagella and cilia become ineffective at rather small size, and amoeboid movement is limited to the size a single cell can attain. Muscles contract by a cellular mechanism basically like that used in amoeboid locomotion—interaction of actin and myosin filaments. Through coordinated contraction of many cells, movement of large individuals becomes possible.

Coelenterates, like parazoans, have only two body layers, an inner endoderm primarily for feeding and an outer ectoderm for protection. Between the endoderm and the ectoderm of coelenterates is the mesoglea, a gelatinous mass that contains connective fibres of collagen and usually some cells. Both layers contain muscle fibres and a two-dimensional web of nerve cells at the base; the endoderm surrounds a central cavity, which ranges from simple to complex in shape and serves as a gut, circulatory system, and sometimes even a skeleton. The cavity is also used for gamete dispersal and waste elimination.
Cleavage of a fertilized egg produces a hollow sphere of flagellated cells (the blastula). Invagination of cells at one or both poles creates a mouthless, solid gastrula; the gastrula is called the planula larva in species in which this stage of development is free-living. The inner, endoderm cells subsequently differentiate to form the lining of the central cavity. The mouth forms once the planula larva has settled. Although the details of early development are different for parazoans and coelenterates, most share a stage in which external flagellated cells invaginate to form the inner layer, which lines the cavity, of these diploblastic (two-layered) animals. This is characteristic of invagination during the development of all animals.
All coelenterates are more or less radially symmetrical. A radial form is equally advantageous for filtering, predatory, or photosynthetic modes of feeding. Tentacles around the circumference can intercept food in all directions.
Bilateria: an organ level of organization
All animals except those in the four phyla mentioned above have bilaterally symmetrical ancestors and contain three body layers (triploblastic) with coalition of tissues into organs. The body plans that are generally recognized are acoelomate, pseudocoelomate, and coelomate.
Acoelomates have no internal fluid-filled body cavity (coelom). Pseudocoelomates have a cavity between the inner (endoderm) and the middle (mesoderm) body layers. Coelomates have a cavity within the mesoderm, which can show one of two types of development: schizocoelous or enterocoelic. Most protostomes show schizocoelous development, in which the mesoderm proliferates from a single cell and divides to form a mass on each side of the body; the coelom arises from a split within each mass. Deuterostomes show enterocoelic pouching, in which the endoderm evaginates and pinches off discrete pouches, the cavities of which become the coelom and the wall the mesoderm. The animals in these major divisions of the Bilateria differ in other fundamental ways, which are detailed below.

Unlike sessile sponges or floating jellyfish, the Bilateria typically move actively in pursuit of food, although many members have further evolved into sessile or radial forms. Directed movement is most efficient if sensory organs are located at the head or forward-moving end of the animal. Organs of locomotion are most efficiently arranged along both sides, a fact that defines the bilateral symmetry; many internal organs are not in fact paired, whereas muscle layers, limbs, and sensory organs almost invariably are. The diffuse nerve net of coelenterates coalesces into definite tracts or bundles, which run posteriorly from the anterior brain to innervate the structures of locomotion.
Acoelomates
Flatworms (phyla Platyhelminthes, Nemertea, and Mesozoa) lack a coelom, although nemerteans have a fluid-filled cavity at their anterior, or head, end, which is used to eject the proboscis rapidly. The lack of a fluid-filled cavity adjacent to the muscles reduces the extent to which the muscles can contract and the force they exert (see below Support and movement). Because most also lack a circulatory system, supplying muscle tissues with fuel and oxygen can be no faster than the rate at which these substances diffuse through solid tissue. Flatworms are thus constrained to be relatively flat and comparatively small; parasitic worms, which do not locomote, can achieve immense lengths (e.g., tapeworms), but they remain very thin. The larger of the free-living flatworms have extensively divided guts, which reach to within a few cells of the muscles, thus compensating for the lack of a circulatory system. Most flatworms have but one opening to the gut. Nemerteans, in addition to a coelom-like housing for their proboscis, have attained a one-way gut and a closed circulatory system. Both increase their ability to move food and oxygen to all parts of the body. Flatworms are considered to be the ancestors of all other Bilateria.
Pseudocoelomates, or aschelminths
The pseudocoelomates include the nematodes, rotifers, gastrotrichs, and introverts. Some members of some other phyla are also, strictly speaking, pseudocoelomate. These four phyla of tiny body size (many species no larger than the bigger protozoans) are placed together in part because they lack mesoderm on the inner side of the body cavity. Consequently, no tissue, muscular or connective, supports the gut within the coelomic fluid. For tiny organisms, this is advantageous for conservation of tissue: there is no reason to evolve or to maintain a tissue that is not functionally important. The inconspicuousness of most of these phyla has led to a slow advancement in understanding their phylogenetic position in the animal kingdom.





